ENTEROBACTERIAS (Última Parte)
“Mantener la buena salud del cuerpo es un deber, por lo que no debemos descuidar el conocimiento y mantener nuestra mente fuerte y clara. El agua rodea la flor de loto, pero no moja sus pétalos.” (Buda)
ENTEROBACTERIAS (Última Parte)

Género Morganella
Morganella,
pertenece a la familia Enterobacteriaceae y hasta el momento, presenta una sola
especie: morganii. Inicialmente, Morganella morganii recibió el nombre de
Proteus morganii, pero por estudios genéticos, se le sacó del género Proteus y
se creó el género Morganella. Este último
género, presenta un INVIC: + + - - , es capaz de hidrolizar la
urea, tiene una movilidad variable a 37° C y produce ácido y gas a partir de la
glucosa. Además, es capaz de reducir los nitratos a nitritos, de fermentar la manosa
y como todos los integrantes de la familia Enterobacteriaceae, es oxidasa
negativo.

Las cepas de
Morganella morganii, crecen bien en los medios de aislamiento primarios como el
agar sangre y el agar MacConkey, no son hemolíticas y usualmente no producen el
fenómeno de "swarming" como Proteus. En épocas recientes, se
propuso la creación de una sub clasificación de este género en dos sub
especies: Morganella morganii ssp. morganii y Morganella morganii ssp. sibonii. M. morganii,
es causa conocida de infecciones del tracto urinario y fuera del tracto
urinario, puede producir diversos tipos de infecciones. Por otra parte, se le
ha relacionado como un patógeno entérico, sin embargo aquí, su papel etiológico
es dudoso. En forma reciente, se documentó un caso de una
corioamnionitis y sepsis asociada, caso complicado con un síndrome de distress
respiratorio, además de infecciones en el recién nacido.

Este agente,
rara vez es causa de infecciones invasivas en personas inmunocompetentes, pero
sí puede ser una causa probable de infecciones nosocomiales en personas inmunocomprometidas. Esta bacteria, presenta una resistencia intrínseca a la
polimixina, ampicilina y a cefalotina y se ha documentado que es capaz de
producir una beta lactamasa inducible o la beta lactamasa de efecto expandido. Todo esto,
claro está, cuando el agente se ve expuesto a un agente inductor y las enzimas
producidas, hidrolizan a las penicilinas y cefalosporinas de efecto expandido.
Generalmente, estas bacterias dejan de producir la enzima cuando el agente
inductor es removido, aún cuando, algunas cepas mutan a un estado de continua
producción de beta lactamasa. Todo esto nos habla de un agente con una alta
resistencia antimicrobiana, tanto natural (intrínseca) como adquirida.

Género
Providencia
Providencia rettgeri (antes Proteus rettgeri) es una bacteria móvil
perteneciente al género Providencia, que junto a Proteus y Morganella forman la
tribu Proteae o Pro. Componente normal de la flora colónica en el hombre, se
diferencia de otras especies de Providencia en que hidroliza rápida y
abundantemente la urea pero no metaboliza a la lactosa como los otros
integrantes de Enterobacterias. Como patógeno humano se ha descripto como causante de infecciones urinarias,
sobre todo en pacientes mayores y con catéteres uretrales, y en infecciones de
heridas en pacientes inmunodeprimidos y quemados. A diferencia de P. stuartii
son muy escasas las publicaciones de bacteriemias causadas por este
microorganismo. En una
revisión bibliográfica en PubMed utilizando los términos ''Bacteriemia y
Providencia rettgeri'' aparecen sólo 2 casos de sepsis fatal en probable
relación a contaminación de hemoderivados y algún caso aislado en grandes series
de bacteriemias.

Por otra
parte, Providencia alcalifaciens es un miembro bastante semejante al anterior y
perteneciente también a la familia Enterobacteriaceae. Hay informes que P. alcalifaciens puede causar diarrea, pero el mecanismo por
el que la causa no es bien conocido. El modelo de condensación de la actina era
diferente del producido por la toxina enteropatogénica de Escherichia coli pero
similar a Shigella flexneri. Se
concluye que algunas algunas subespecies de P. alcalifaciens son los
enteropatogénicos y que ellos causarían los cuadros de diarrea invadiendo el
epitelio de la mucosa intestinal.

Género
Yersinia
Este género
comprende varias especies entre ellas Yersinia pestis, agente de la
Peste Negra o Plaga Bubónica o Neumónica, comúnmente llamada la
Muerte Negra, enfermedad de los roedores, transmitida ocasionalmente al
hombre por las pulgas, con pandemias históricas desde el siglo VI, donde mató a
un tercio de la población en Europa. Luego de la Edad Media ha habido
brotes en diversas partes del mundo, sobre todo en relación con las guerras. Se han
denunciado en 1995 y comienzos de 1996 casos en India, Madagascar y otros
países africanos, en Brasil y Perú. Y. pestis es endémica en algunas regiones
tales como Irán y el oeste de Estados Unidos. Otra especie, Y. enterocolitica es muy ubiquitaria,
se ingiere con agua o alimentos contaminados. Algunos biotipos están
relacionados con enterocolitis en el hombre. Raramente presenta infecciones
sistémicas. Sin embargo, las bacterias atraviesan con frecuencia la mucosa y se
multiplican en los nódulos linfáticos mesentéricos. Debido a los intensos
dolores abdominales el cuadro puede confundirse con apendicitis. Ocasionalmente
puede haber una artritis reactiva 2 a 6 semanas luego de la
infección. Esto se ve frecuentemente en pacientes con antígeno HLA-B27 de
histocompatibilidad. Cepas virulentas de las 2 especies poseen al menos un gran
plásmido de virulencia.
Y.
enterocolitica es una bacteria pequeña de forma redonda y Gram-negativa,
la cual es aislada frecuentemente de los especímenes clínicos tales como las
heridas, las heces fecales, el esputo o las glándulas linfáticas mesentéricas.
Sin embargo, no forma parte normal de la flora humana. Por otro lado, Y.
pseudotuberculosis ha sido aislada del apéndice infectado en los humanos. Ambos
organismos han sido aislados frecuentemente de los animales, tales como los
cerdos, las aves, los castores, los gatos y los perros. Solamente la bacteria
Y. enterocolitica se ha encontrado en muestras ambientales de lagunas y lagos,
y en alimentos como la carne, los helados y la leche. La mayoría de los
organismos aislados no han sido catalogados como patógenos. El nombre de la enfermedad causada por
este microorganismo es yersiniosis.

Existen tres especies patogénicas dentro del género Yersinia, pero sólo Y.
enterocolitica y Y. pseudotuberculosis causan gastroenteritis. Hasta el
momento, se han reportado muy pocos casos de brotes causados por Y.
pseudotuberculosis, como por ejemplo las presentadas en Japón, donde se
reportaron infecciones en humanos transmitidas por alimentos y por aguas
contaminadas. La
yersiniosis se caracteriza generalmente por síntomas tales como la
gastroenteritis con diarrea y/o con vómito; sin embargo, la fiebre y el dolor
abdominal son los síntomas que la definen. Las infecciones causadas por
Yersinia son similares a la apendicitis y a la linfadenitis mesentérica, pero
esta bacteria también puede causar infecciones en otras áreas como en las
heridas, en las articulaciones y en el tracto urinario.

La dosis infecciosa es desconocida. La aparición de la enfermedad se da entre
las 24 y 48 horas después de la ingestión, la cual es la ruta usual de
infección (los alimentos y las bebidas son los vehiculos de transmisión). El diagnóstico de la yersiniosis se
inicia con el aislamiento del microorganismo a partir de las heces fecales, de
la sangre, o del vómito de la víctima, y a veces la muestra es tomada durante
la apendicectomía. La confirmación se realiza con el aislamiento y la posterior
identificación bioquímica y serológica de Y. enterocolitica, provenientes tanto
del hospedero humano como del alimento ingerido. Se ha reportado que la diarrea
ocurre en el 80% de los casos; y que los síntomas más confiables son el dolor
abdominal y la fiebre. Dada la dificultad para el aislamiento de yersiniae de las heces fecales,
muchos países dependen de la serología. Para ello, a los pacientes críticos y
convalecientes se les realiza un análisis en busca del serotipo de Yersinia
spp. sospechoso.

Las cepas de
Y. enterocolitica se pueden encontrar en las carnes (de cerdo, la de carne de
vaca, la de cordero, etc.), en las ostras, el pescado, y la leche cruda. La
causa exacta de la contaminación de los alimentos es desconocida. Sin embargo,
la prevalencia de este organismo en el suelo y el agua, así como también en
ciertos animales como los castores, los cerdos, los conejos y las ardillas,
ofrece grandes oportunidades para que este organismo ingrese a la cadena
alimentaria. La falta de higiene de los manipuladores de alimentos y las
técnicas de esterilización inadecuadas, además del almacenamiento inapropiado,
son también importantes fuentes que contribuyen a la contaminación. Yersinia es sensible al calor y
como resultado morirá durante el calentamiento (mayor a 70°C). L as
principales causas de infección son el consumo de los alimentos crudos o
parcialmente cocidos y así como la contaminación cruzada, que ocurre cuando los
productos cocidos entran en contacto con los materiales crudos o contaminados
(tablas para cortar). Por esta razón, la cocción adecuada y la higiene en el
manejo de los alimentos pueden prevenir las infecciones causadas por Yersinia
en una gran medida. Las
poblaciones más susceptibles a la enfermedad principal y a sus posibles
complicaciones son las más jóvenes, también los débiles, los ancianos y las
personas que se encuentren bajo terapias inmunosupresoras.

El nombre
del género Yersinia viene dado por su descubridor, en 1894, Alexandre Yersin,
bacteriólogo franco-suizo del Instituto Pasteur. Originalmente, este
microorganismo fue denominado Pasteurella pestis, pero en el año 1967 fue
renombrado como Yersinia pestis en honor a su descubridor. Yersinia pestis es un agente infeccioso que ha sido directamente
responsable de más muertes humanas que cualquier otra enfermedad infecciosa,
salvo la malaria. Ha originado diversas pandemias a lo largo de la historia,
entre las que cabe destacar: la Plaga de Justiniano (541-542 dC), que
asoló Asia, el norte de África, Arabia y parte de Europa; la Peste
Negra (1347-1351 dC), que acabó con la vida de un tercio de la población
de Europa; y la Tercera Pandemia (1855-1959), que comenzó en China e
India y terminó por extenderse por el resto de Asia, África y América.

El papel de
Y. pestis en la Peste Negra se ha debatido entre los historiadores;
algunos han sugerido que la Peste Negra se propagó muy rápido para
haber sido causada por Y. pestis. Se ha encontrado ADN de Y. pestis en los
dientes de aquellos que murieron de Peste Negra pero, sin embargo, cadáveres
medievales que murieron de otras causas no dieron positivo a Y. pestis. Esto
sugiere que Y. pestis fue un factor que contribuyó a las plagas europeas, pero
probablemente no el único. Es posible que las presiones selectivas inducidas
por la plaga puedan haber cambiado el modo en que el patógeno se manifiesta en
humanos, seleccionándose en contra de individuos o poblaciones que eran más
susceptibles.
- Yersinia
pestis, agente de la denominada peste bubónica o peste negra, que asoló
Europa a mediados del siglo XIV. Esta especie presenta tres subespecies:
Y. pestis antiqua, Y pestis medievalis y Y. pestis orientalis.
- Yersinia
pseudotuberculosis, agente de una enfermedad parecida a la tuberculosis
que afecta a los nódulos linfáticos en animales y raramente en humanos.
- Yersinia
enterocolitica, agente de ciertas infecciones intestinales (a veces
también sistémicas) en humanos y animales.

El género Yersinia pertenece a las bacterias Gram-negativas aeróbicas
facultativas con metabolismo fermentativo. Es un cocobacilo de tinción bipolar
similar a otras Enterobacterias. Durante el proceso infeccioso genera
viscosidad antifagocítica. El organismo presenta motilidad cuando es aislado,
pero pierde esta capacidad mientras permanece en el mamífero huésped. La peste es una enfermedad natural de
los roedores, siendo las ratas el principal reservorio de la enfermedad. Tras
ser infectadas, la mayoría de las ratas mueren, pero un pequeño porcentaje
consigue sobrevivir, quedando como una fuente de Y. pestis. Las ratas son infectadas a través de un vector, que en este caso es la pulga de
rata (Xenopssylla cheopis). La pulga chupa la sangre de un animal infectado e
ingiere a la vez células de Y. pestis, las cuales se multiplican en el
intestino de la pulga y serán transmitidas a otra rata en la siguiente picadura
de la pulga. La
enfermedad se irá extendiendo de forma que la mortalidad entre las ratas se
hace tan elevada que la pulga busca nuevos hospedadores, entre los que se
encuentra el hombre.

A continuación se exponen los tres tipos de infecciones a las que puede dar lugar Yersinia pestis:
Peste
bubónica: En el tipo de infección más común. Una vez que las células
bacterianas han sido introducidas mediante la picadura de una pulga dentro de
un ser humano, las células bacterianas se desplazan por el torrente sanguíneo
hasta los nódulos linfáticos donde generan pequeñas hinchazones denominadas
bubones, que están llenos de partículas bacterianas. La cápsula viscosa que
rodea a las células de Y. pestis evita que estas sean fagocitadas por los
macrófagos. En poco tiempo, los nódulos linfáticos periféricos se ven invadidos por bubones
secundarios, hasta que se rompen y las células pasan de nuevo al torrente
circulatorio, pero ahora en un número mucho más elevado, lo que causa una
septicemia generalizada. En este
estado, se producen múltiples hemorragias que dan lugar a manchas negras sobre
la piel, procesos de gangrena en los extremos distales de las extremidades,
fuerte dolor en nódulos linfáticos, postración, shock y delirio. Si la peste no
es tratada antes del estado septicémico, la muerte sobreviene al cabo de 3-5
días.

Peste Neumónica: Este tipo de infección se produce cuando las células de
Y. pestis son inhaladas directamente, o bien llegan a los pulmones durante la
peste bubónica. La infección suele transcurrir sin síntomas hasta los dos
últimos días del proceso infectivo, en los cuales se emiten gran cantidad de
esputos con sangre. En ausencia de tratamiento la muerte sobreviene en 2-3
días.
Peste septicémica: Esta infección implica una rápida dispersión de Y.
pestis por todo el cuerpo, a través del torrente circulatorio, sin tiempo para
que se formen los bubones. La muerte suele sobrevenir en 1 día, por lo que
habitualmente no da tiempo a ser diagnosticada.

Las células de Y. pestis producen ciertas moléculas antigénicas que contribuyen
en mayor o menor medida al proceso de la enfermedad.
- En
la pared bacteriana se encuentran unos complejos de proteína-lipoproteína,
denominados antígenos VW y F1, cuya función es evitar la fagocitosis por
parte de los macrófagos y otras células fagocíticas.
- Las
cepas virulentas de Y. pestis producen y secretan una exotoxina denominada
toxina murina (debido a su extrema toxicidad para el ratón), cuyo modo de
acción se basa en inhibir la respiración, bloqueando las reacciones de
transporte de electrones en las mitocondrias, a nivel de la coenzima Q.
Aunque no está claro que la toxina murina esté implicada en la patogénesis
de la peste humana, los síntomas que produce en el ratón son similares a
los que produce en humanos.
- Y.
pestis también produce una endotoxina altamente inmunogénica que
puede dar lugar a una potente respuesta inmune en humanos.
Y. pestis presenta resistencia natural a la penicilina, pero la mayoría de las cepas son sensibles a la estreptomicina, el cloranfenicol y las tetraciclinas. Actualmente hay ciertas evidencias de la sensibilidad de Y. pestis a gentamicina y doxiciclina. Si el tratamiento se inicia rápidamente, la mortalidad de la peste bubónica puede reducirse hasta el 1-5% de los infectados. La peste neumónica y septicémica también puede tratarse, pero suelen progresar tan rápidamente que los antibióticos siempre llegan tarde.
Bacilos
Gram (-) no fermentadores
Este gran
grupo de bacilos Gram negativos incluye a gérmenes pertenecientes a diferentes
familias y otros géneros de incierta clasificación. Pseudomonas,
Flavobacterium, Alcaligenes, Acinetobacter, son algunos de ellos, en general
desprovistos de grandes atributos de virulencia demostrables, no producen
enfermedad en el individuo sano pero pueden comportarse como oportunistas en
enfermos inmunodeprimidos. De las
numerosas especies de Pseudomonas descritas sólo unas pocas tienen importancia
en patología humana. Pseudomonas mallei y P.pseudomallei causan enfermedad
severa en el hombre pero se aíslan raramente en el Hemisferio Occidental. Por
otra parte P. cepacia es un oportunista poco frecuentemente asociado con
enfermedad en el hombre.

Nos
referiremos en particular a la especie Pseudomona aeruginosa por su frecuencia
en patología humana y estar mejor estudiada que otros. Es un microorganismo
versátil, ampliamente distribuido en el suelo, agua, plantas e intestino de
animales. Puede causar enfermedad en el hombre, ciertos animales, plantas e
insectos. El agua
contaminada puede ser una fuente de infección para el hombre. Es susceptible a
la desecación, pero sus habilidades metabólicas le permiten sobrevivir y
multiplicarse en líquidos y ambientes húmedos de los hospitales. Sus
requerimientos nutricionales son variados, se ha aislado P. aeruginosa de aguas
termales, e incluso de soluciones desinfectantes en el hospital. Las
infecciones humanas están la mayoría restringidas a los pacientes
hospitalizados que adquieren el microorganismo de fuentes ambientales
(infección exógena) por contacto con vectores humanos o inanimados. P.
aeruginosa desarrolla bien en medios simples, utilizándose para su aislamiento
los medios de cultivo de uso corriente en el laboratorio clínico.

La identificación de cepas de P. aeruginosa típicamente productoras de pigmento no es difícil, pero las cepas no pigmentadas pueden presentar un problema. La mayoría se identifican por la producción de un pigmento, pyocyanina (Bacilo piociánico), soluble en agua, azul, no fluorescente. P. aeruginosa produce además otro pigmento, pyoverdina, soluble en agua, verdeamarillento, fluorescente; otras especies del género Pseudomonas también producen pyoverdina. Otros pigmentos, menos frecuentes pueden ser producidos por P. aeruginosa. La morfología colonial y el olor frutado de aminoacetofenona son elementos de una identificación sencilla, y aunque existen caracteres de identificación confirmatorios, son de uso poco corriente. Son bacilos Gram negativos, rectos o ligeramente curvos, móviles, con un solo flagelo polar. Oxidasa y catalasa positivas, aerobias estrictas, no fermentan glucosa, utilizan diversos azúcares oxidativamente con producción de ácido. Uno de los caracteres más constantes es su capacidad de desarrollar a 42ºC. Producen varias enzimas, proteasas, lipasas, lecitinasas.

Las defensas
inespecíficas del huésped son en general suficientes para prevenir la infección
por P. aeruginosa, pero brechas en esta barrera permiten a P. aeruginosa
invadir y causar infecciones de diversa gravedad. Producen el 10% de las
infecciones nosocomiales, infectan heridas y quemaduras y causan infecciones
pulmonares, sobre todo neumonía nosocomial e infecciones respiratorias en
pacientes con fibrosis quística. La fibrosis quística es una enfermedad genética asociada a un defecto en la
secreción de cloro, caracterizada por la producción de mucina con una
alteración de su composición iónica, inusualmente espesa. Esto lleva a una
menor eficiencia de la mucina para limpiar las bacterias del pulmón y las vías
aéreas y puede impedir el movimiento de las células fagocíticas. Estos hechos
explican la susceptibilidad de los pacientes con fibrosis quística a la
colonización con P. aeruginosa. Si los enfermos son tratados los síntomas
pueden desaparecer pero las bacterias permanecen, presentando infecciones
recurrentes. Las condiciones del paciente se ven agravadas con la infección a
P. aeruginosa por las dificultades terapéuticas que se plantean debido a su
alta resistencia a los antimicrobianos.

P.
aeruginosa posee los mismos tipos de factores de virulencia que otras
bacterias capaces de causar enfermedad en el hombre inmunocompetente. Pero algo
interesante es ¿por qué P. aeruginosa no es un patógeno franco y es sólo capaz
de producir infecciones oportunistas? Es probable
que P. aeruginosa sea ineficiente en su habilidad para llevar a cabo los
primeros pasos de la infección; puede colonizar pero no invadir piel y mucosas
sanas y tampoco dar infecciones persistentes con producción concomitante de
factores tóxicos que dañen los tejidos del huésped. Adhesinas. Produce dos tipos de adhesinas proteicas, pili y adhesinas no pili.
Los pili son pili tipo 4 similares a los de N. gonorrhoeae y se parecen también
a los pili Tcp de V. cholerae. Permiten a la bacteria adherirse a las células
epiteliales, preferentemente a receptores asialo-GM1. P. aeruginosa produce una
neuraminidasa que saca los residuos de ácido siálico de GM1, creando sitios de
unión para la pilina.

Por otra
parte, P. aeruginosa es capaz de unirse a la mucina y lo hace por medio de las
adhesinas no pili. Además del gen que codifica para la proteína estructural del
pili otros genes codifican proteínas ensambladoras y reguladoras. Exoenzima S.
Es una enzima excretada que puede actuar como exotoxina. Tiene actividad de
ADPribosilación como otras toxinas, pero aplicada en forma exógena no daña las
células del huésped. Al igual que la toxina colérica intervienen proteínas de las células del
huésped en la activación de la toxina para lograr su máxima actividad. Se
sostiene que actuaría dificultando la acción de los fagocitos lo que
facilitaría la sobrevida de P. aeruginosa en el torrente sanguíneo y órganos.
En el pulmón actuaría inhibiendo la muerte intrafagocítica de las bacterias y
promoviendo la infiltración fagocítica en el área. También puede presentar
efecto tóxico directo en los pulmones.

Exotoxina A. Esta exotoxina tiene el mismo mecanismo que la toxina diftérica. Es una toxina A-B con tres unidades funcionales:
- dominio R
(región de unión al receptor celular),
- dominio T
(región que media la translocación de la porción enzimática al interior de la
célula),
- dominio C
(región catalítica).
Los dominios
R y T se localizan en la cadena B y el dominio C en la cadena A. La cadena A es
enzimáticamente activa por ADPribosilación del factor de elongación 2 (EF-2) de
la síntesis proteica, que lo vuelve inactivo. Su receptor es una glicoproteína
de las células del hospedero. La mayoría de los aislamientos clínicos la
producen, y actuaría produciendo daño en los tejidos y disminuyendo la
actividad de los fagocitos. Elastasas.
Elastina es el 30% de las proteínas del tejido pulmonar. Está también presente
en la pared de los vasos sanguíneos. Es responsable de las propiedades
elásticas de estos órganos que se expanden y contraen. P. aeruginosa tiene
actividad elastolítica, produce dos enzimas que actuarían concertadamente: LasA
y LasB. LasA actuaría clivando la elastina y permitiendo la acción de LasB, que
es una zinc metaloproteasa, uno de cuyos sustratos es la elastina.

Estas
enzimas actuarían en las etapas tempranas de la enfermedad, por daño directo de
los tejidos pero no en infecciones crónicas, debido a la presencia de
anticuerpos antielastasas. También pueden intervenir degradando componentes del
complemento e inhibidores de 1 proteinasa (inhibe el daño de los tejidos por
las proteasas de los polimorfonucleares (PMNs)). En las infecciones crónicas,
altos niveles de anticuerpos producidos pueden llevar a la formación de
complejos inmunes y su depósito en el pulmón activar complemento y atraer PMNs.
Los PMNs producen su propia elastasa, más potente que LasA-LasB. Pequeñas
cantidades de LasA pueden facilitar la degradación de la elastina pulmonar
causada por la elastasa de los PMNs. Otras enzimas extracelulares. Produce
varias enzimas además de las mencionadas. Una lipasa alcalina y dos
fosfolipasas, no bien estudiadas. Por otra parte, pyocianina puede funcionar
como factor de virulencia. Puede dañar el tejido endotelial in vitro, lo que
sugiere una acción in vivo.

Un atributo
de virulencia muy interesante es la producción de alginato. Es un polímero de
ácido mannurónico y gulurónico que forma un gel viscoso alrededor de la
bacteria. Las colonias que lo producen tienen aspecto mucoide. Para las
bacterias marinas esto es un atributo importante para su supervivencia. P.
aeruginosa ha adaptado esto a su supervivencia en el pulmón. En medios de
cultivo ricos pierde esta propiedad. Esta capa
que rodea a la bacteria y a las colonias de bacterias en el pulmón puede actuar
como adhesina y probablemente previene la ingestión fagocítica de la bacteria.
Los genes que intervienen en su codificación están agrupados en un sector del
cromosoma y organizados en un operón, poseen un sistema de regulación
extremadamente complejo. El LPS
también varía durante la transición mucoide – no mucoide. En cepas no mucoides
el antígeno O del LPS tiene cadenas largas y carga negativa mientras que las
cepas mucoides tienen cadenas más cortas y una composición de azúcares que lo
hacen mucho más neutro; esto sería importante en la alta resistencia a algunos
antibióticos que presenta P. aeruginosa, situación problemática en pacientes
internados, pero dramática en los pacientes con fibrosis quística, que muchas
veces presentan infecciones por P. aeruginosa resistente a todos los
antibióticos disponibles.

Género Hafnia
A éste último grupo de enterobacterias me
gusta llamarlas "las figuritas difíciles del álbum" pues son raras de
encontrar y de aislar. Hafnia alvei es un bacilo gram negativo, única
especie del género Hafnia, de la familia Enterobacteriaceae, y que durante
mucho tiempo fue incluida en el género Enterobacter, hasta el desarrollo de las
técnicas de biología molecular, que determinaron su separación, con definición
de un nuevo género. Se encuentra ampliamente disperso en la naturaleza, en
lugares como aguas residuales, suelo, oro – faringe humana, intestino delgado y
heces de personas y animales siendo habitualmente un microorganismo
colonizador. Se trata de un microorganismo facultativo,
usualmente no patógeno, que se ha aislado de cultivos orgánicos (faringe,
bronquio, sangre, orina y heces), produciendo infecciones en la gran mayoría de
los casos, en pacientes con enfermedad crónica de base, inmunodeprimidos y en
relación con estancias hospitalarias y utilización de antibioticoterapia de
amplio espectro.

En los casos descriptos en la literatura el
aislamiento de Hafnia alvei se asocia a flora polimicrobiana, siendo difícil
establecer su implicación como agente causal. En la bibliografía revisada
(MEDLINE de los últimos 10 años) se han descrito casos de gastroenteritis,
enterocolitis necrotizante, colecistitis piógena, peritonitis, meningitis,
infecciones del tracto urinario, abscesos cutáneos, endoftalmitis,
bacteriemias, endocarditis, neumonía, empiema e infecciones de heridas en
enfermos hospitalizados. H. alvei es un bacilo cuyo nombre es
enterobacter hafniae y puede ser responsable de infecciones graves adquiridas
en la comunidad o en el hospital, constituyendo Enterobacter y Hafnia, agentes
de infección nosocomial en el 5% de los casos. En adultos se considera una
causa excepcional de bacteremia, neumonía, infección nosocomial de tejidos
blandos, colonización de catéteres centrales, infección postransplante,
endocarditis protésica, septicemia con shock y coagulación intravascular
diseminada. Esto es así porque los sujetos con infección
por H. alvei, excepto aquéllos con afectación intestinal, suelen tener una
enfermedad grave de base, enfermedad pulmonar obstructiva crónica, diabetes
mellitus, insuficiencia renal crónica, cáncer o una serología VIH positiva.

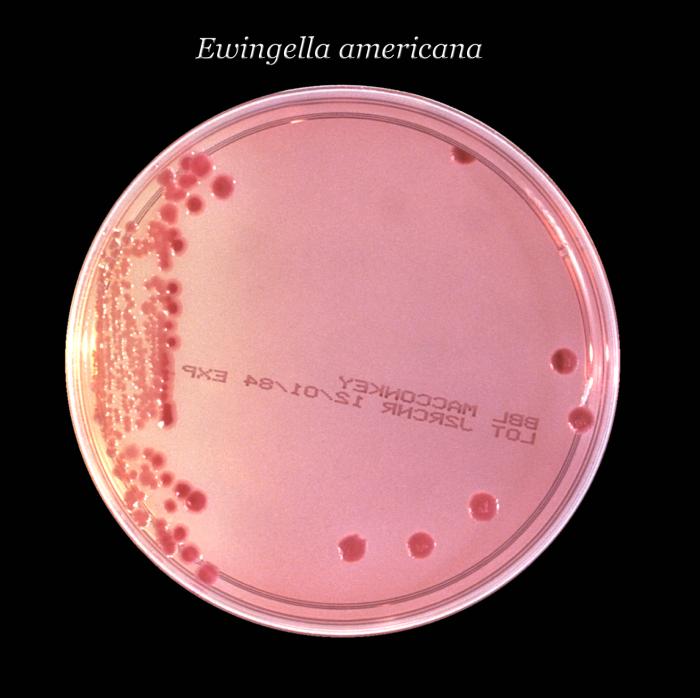
Género Ewingella
Ewingella, previamente conocido como Grupo Entérico 40, es un
nuevo género en la familia de Enterobacteriaceae y se describió primero por
Grimont y colaboradores en 1983. Este
género incluye sólo una especie, la
Ewingella americana.
Este microorganismo raramente se encuentra en las muestras clínicas humanas. La
fuente más común ha sido sangre, pero también se ha aislado del esputo,
orina y heridas. En la mayoría de los casos, la importancia de
la patogenia de este organismo, aunque sugestivo, no se ha establecido.
Recientemente, informes esparcidos de infecciones debidas a E. americana han
aparecido en la literatura. Ahora es más frecuente observar su desarrollo
en peritonitis causado por E. americana en pacientes con fases finales de
enfermedad renal que sufren la diálisis peritoneal ambulante continua desde
hace más de 12 años. La causa primaria de fracaso renal era la enfermedad del
riñón poliquístico.
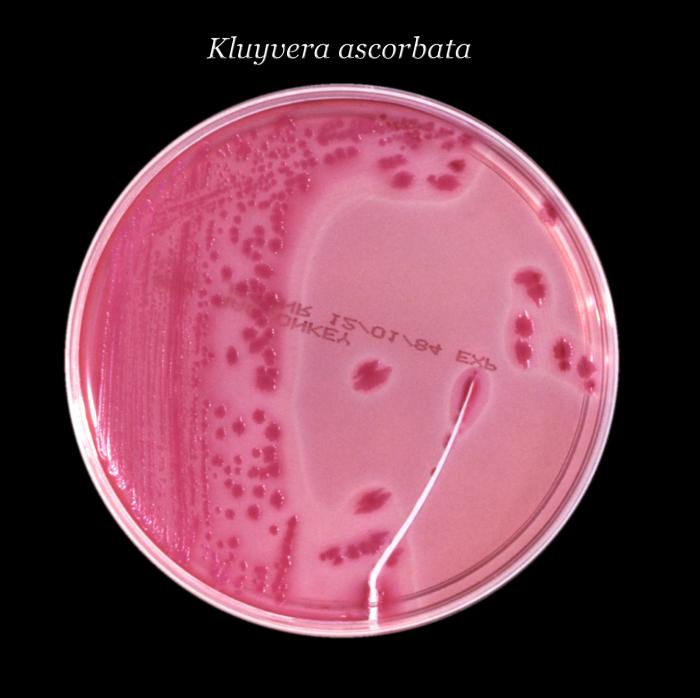
Género Kluyvera
Kluyvera corresponde
a un nuevo género llamado: Grupo Entérico 8. Se divide en tres grupos:
• Kluyvera ascorbata, que sería la especie
tipo y que se aísla preferentemente de especímenes clínicos, es incapaz de
crecer a 5º C, e in vitro es inhibida por cefalotina y carbenicilina. Es test
de ascorbato positivo a diferencia de la segunda especie,
• Kluyvera cryocrescens que es negativa ante este test y que
se aísla más del medio ambiente que del paciente.
• Finalmente existe un grupo heterogéneo, sin
nombre específico llamado Kluyvera especie grupo tres, distinto genéticamente
de los anteriores.
Las cepas de Kluyvera tienen las mismas
características de la familia Enterobacteriaceae a la cual pertenecen, son
bacilos Gram (-) con flagelos perítricos lo que la hacen móviles, catalasa (+)
y oxidasa (-), crecen en agar de Macconkey, fermentan la glucosa y son indol
(+), rojo metilo (+). Vogues Proskauer (-), citrato (+), H2S (-),
ureasa (-), fenilalanina deaminasa (-) y arginina dihidrolasa (-), lisina (+) y
ornitina descarboxilasa (+). Es difícil diferenciarlas de Escherichia
coli. Según resultados a través del Microscan, pudimos apreciar que es una
colonia más seca y rugosa y con un fenotipo de mayor resistencia a
antimicrobianos que E. coli. Todavía se requiere mayor información
para que podamos adjudicarle un significado clínico a este género.
Cedecea davisae
es un bacilo gramnegativo, que pertenece a la familia de las
Enterobacteriaceae; aunque tiene diferencias fenotípicas con los otros
microorganismos de esta familia de bacterias, tiene grandes similitudes con la
Serratia. Es lipasa positiva, siendo resistente a la
acción antimicrobiana de algunas cefalosporinas y de la colistina. Se han
comunicado aislamientos por este infrecuente microorganismo en esputo, tracto
gastrointestinal, heridas de las manos, escroto y ojos y también se ha aislado
en algunos casos de bacteriemias. No se ha encontrado a
la fecha ningún caso de absceso de puerta de entrada de laparoscopia en el que
se haya aislado este microorganismo, pero no es de extrañar que sea un
oportunista en éste tipo de técnicas quirúrgicas. El nombre de Cedecea fue propuesto
recientemente para un único nuevo género en el ancho mundo de las
Enterobacteriaceae, previamente conocido como Grupo Entérico 15. Los extendidos
de Cedecea se parecen a los de Serratia porque ellos son lipasas positivas y
resistentes al cephalothin y colistin. Sin embargo, Serratia difiere de Cedecea al
no hidrolizar la gelatina y el ADN. Dos especies se nombraron originalmente en
Cedecea: C. davisae y C. lapagei, pero la hibridación de ADN indicó que otro
grupo era distinto y debe ser considerado entonces como un tercero adicional.

Género Moellerella
El nombre de Moellerella wisconsensis se
propone previamente para un grupo de la familia Enterobacteriaceae llamado los
Grupos Entéricos 46. La especie wisconsensis se acuñó porque se aislaron seis
de las nueve especies reconocidas en Wisconsin (USA). Estas
especies de wisconsensis eran negativas para: la producción del indol, Voges –
Proskauer, producción de SH2, urea, deaminasa de la felilalanina,
lisina y decarboxilasa de la ornitina, dihidrolasa de la arginina, la
producción de gas de la
D –
glucosa, la producción ácida de la trealosa y la motilidad. En cambio, eran positivas para: el rojo de
metilo, citrato (Simmons), producción ácida de la lactosa y rafinosa y
resistente al colistin. La información clínica sobre un caso de
un paciente internado encontrado en USA puede a priori ser asociado con un caso
de diarrea del mismo no por su patogenicidad sino por su oportunismo. En el agar de MacConkey, las colonias de M.
wisconsensis eran luminosas y rojas con la bilis precipitada alrededor de ellos
y así eran indistinguible de las colonias de Escherichia coli. Los estudios
futuros deben enfocar en el aislamiento de este nuevo organismo y su relación a
la enfermedad humana, por sobre todo a la trasmisión por los alimentos (ETA).

Género Rahnella
Entre los cambios que ha experimentado el
trabajo con bacterias y hongos tenemos los adelantos en la identificación, que
ha pasa o rápidamente manual a automatizada, lo que ha permitido crear una
serie de bases de datos con las cuales se aumentan las posibilidades de
identificación correcta de un organismo. La
primera de esas bases de datos es la utilizada por el sistema API de la
Casa BioMerieux , la cual ha mejorado e
introducido una serie de nuevos organismos. Esta misma base de datos se utiliza
en los sistemas automatizados como el Vitek de la misma casa comercial y otras
bases de datos de características similares se emplean en sistemas como el
MicroScan de la
Casa Dade. Unido a esta mejoría en la identificación de
los microorganismos y como un paso más adelante, está el hecho de que el futuro
en la clasificación de los microorganismos parece dirigirse al uso de los
métodos moleculares, ya sea PCR o hibridización. Este
futuro ya casi está entre nosotros y técnicas como el ribotipeo o técnicas para
la detección de genes de resistencia bacterianos ya son de uso en algunos
laboratorios de investigación.

Todo esto ha permitido ampliar la gama de
organismos que los laboratorios clínicos pueden identificar, entre ellos los de
muy baja frecuencia de aislamiento a nivel mundial, como es el caso de la
Rahnella aquatilis
que es un raro bacilo Gram negativo que pertenece a la familia Enterobacteriaceae
y fue aislado por primera vez en 1979. Ha sido
aislado principalmente de agua fresca y los aislamientos clínicos son sumamente
escasos. Ha sido encontrado en el catéter de un
paciente con bacteremia posterior a un transplante de médula ósea, en orinas de
pacientes que han recibido un transplante renal, en heridas quirúrgicas
infectadas, en sangre, en muestras respiratorias y en heces. Algunos de estos
pacientes estaban infectados con el virus de la inmunodeficiencia adquirida. Es
importante mencionar que dada la rareza de los aislamientos de Rahnella
aquatilis en muestras humanas, no es posible establecer, con algún grado de
certeza, su verdadera capacidad patogénica.
"SOMOS LO QUE HACEMOS REPETIDAMENTE. EXCELENCIA, POR LO TANTO, NO ES UN ACTO SINO UN HÁBITO"
ARISTOTELES
LEGALES: El autor no asume responsabilidad alguna por la descarga, copia, distribución, modificación o alteración de los contenidos publicados, sean propios del mismo o de terceros, los cuales pudieren estar protegidos por Copyright, Derechos de Propiedad Intelectual, Derechos de Autor, o relacionados. La Bibliografía del tema expuesto y el crédito fotográfico está en poder del Autor y no se publica dada su extensión, pero se enviará por mail al interesado que la solicitare debidamente fundamentada.



Comentarios
Publicar un comentario