LAS ENTEROBACTERIAS (Parte 2)
Género Escherichia

Como ya fue mencionado E.coli puede
integrar la flora normal, causar diarrea, infección urinaria, meningitis, etc.
Pero una cepa que causa diarrea no causara infección urinaria ni meningitis. La
versatilidad de este microorganismo esta dado porque E.coli ha adquirido conjuntos
diferentes de genes de virulencia. Es el indicador sanitario por excelencia y es un excelente ejemplo de que el poseer un
conjunto de genes es lo que hace que una bacteria sea patógena y no la
designación de género o especie.
Se ha propuesto para E. coli agente de diarrea, una clasificación
de acuerdo a sus mecanismos de virulencia, los llamados virotipos. Aunque
arbitraria, ésta clasificación es muy útil.

Se describen 5 virotipos:
1. E. coli enterotoxigenico (ETEC)
2.
E. coli enteroagregativo
(EAggEC)
3.
E. coli enteropatogénico
(EPEC)
4.
E. coli enterohemorrágico
(EHEC)
5.
E. coli enteroinvasor
(EIEC)
1. E. coli enterotoxigénico (ETEC)
Se parece mucho a V. cholerae, adhiere a la mucosa del intestino delgado, no la
invade, elabora toxinas que causan diarrea. No hay cambios histológicos en las
células de la mucosa y muy poca inflamación. Clínicamente hay diarrea acuosa,
vómitos y se puede acompañar de fiebre. Es la llamada infección no inflamatoria
del intestino delgado.

Para adherirse a las células de la mucosa
ETEC produce diversos tipos de pili. Un tipo de ellos los llamados factores antigénicos de colonización I
y II (CFA/I yCFA/II) parecen contribuir fuertemente a la colonización por estos
microorganismos. Están aun en estudio los receptores para estas adhesinas pero
se piensa que son glicoproteínas. Los genes que codifican para CFA están frecuentemente
localizados en plásmidos.
La diarrea producida por cepas de ETEC es causada
por la acción de dos diferentes toxinas: toxina termolábil (LT) y toxina
termoestable (ST). Hay dos LT y su estructura y mecanismo de acción es el de la
toxina colérica. Tienen diferencias en la excreción de la célula bacteriana y
en la regulación genética de su síntesis. ST es una familia de pequeñas
toxinas. Los genes que codifican para LT y ST son
portados por plásmidos. A menudo el mismo plásmido lleva los genes de las
adhesinas y toxinas.
2. E.coli enteroagregativo (EAggEC)
Son agentes de diarrea persistente. Las cepas de EAggEC se parecen a
ETEC en que se unen a las células intestinales, no son invasivas y no causan modificaciones
histológicas en las células de la mucosa. Difieren de ETEC en que no adhieren
en forma uniforme sino que lo hacen en pequeños agregados.

Estas cepas poseen unas estructuras
fibrilares muy delgadas que se presumen son los pili de adherencia. Aunque es
posible que estos pili promuevan la adherencia de estas bacterias entre sí, más
que la adherencia a la célula del hospedero.
Poseen una forma de adherirse en agregados,
produciendo una toxina similar a ST llamada EAST (ST enteroagregativa). Otra toxina producida por EAggEC es
una toxina muy similar a una hemolisina producida por cepas de E.coli que
causan infecciones urinarias. Esta toxina no hidroliza eritrocitos pero produce
poros en las membranas celulares del hospedero.
3. E.coli enteropatógeno (EPEC)
Es causal de diarrea severa y de gran trascendencia en países subdesarrollados.
EPEC exhibe un patrón de adherencia en parches, pero no forma el mismo tipo de
agregados que EAggEC. A diferencia de las anteriores la adherencia de EPEC
produce alteraciones importantes en la ultraestructura de las células del
huésped. Las células a microvellosidades donde EPEC no se encuentra y estas desaparecen
en el sitio donde la bacteria está adherida.

Este fenómeno se refiere como de unión y
borramiento y es el resultado de un reordenamiento de actina en la vecindad de
la bacteria adherida. EPEC es más invasora que las anteriores y se produce una reacción
inflamatoria. La diarrea producida por EPEC es una
enfermedad más compleja y se piensa que sucede en tres etapas. En un inicio,
hay una asociación de la célula bacteriana a la célula del hospedero llamada unión no íntima, mediada por pili.
Este pili llamado Bfp parece no
ser la única adhesina de EPEC.
Posteriormente se producen señales hacia el
interior de la célula asociadas con activación de enzimas celulares y aumento
de los niveles de Ca++ intracelular, probablemente debido a fosforilización de proteínas
del citoesqueleto y la activación de enzimas despolimerizantes de actina. La
bacteria se asocia entonces más próximamente con la célula del hospedero (unión íntima) produciéndose un reagrupamiento
de actina en la vecindad de la superficie celular. Histológicamente la
deformación de algunas microvellosidades y destrucción de otras se acompaña de
la formación de estructuras similares a pedestales en la célula por debajo del
sitio de adherencia de la bacteria. Estos pedestales son fibras densas de
actina. La unión íntima esta mediada por una proteína de membrana externa
llamada intimina.

Seguramente otras proteínas aun no
identificadas se encuentran también involucradas en este proceso. Algunas
bacterias son posteriormente internalizadas dentro de vesículas fagocíticas.
Muchos de los genes que codifican estos factores han sido localizados en plásmidos.
4. E.coli enterohemorrágico (EHEC)
Se ha reconocido recientemente a EHEC como responsable de cuadros
graves. Estas cepas causan una enfermedad que clínicamente se parece a la
disentería producida por Shigella, aunque probablemente no invade las células de la
mucosa. La enfermedad producida por EHEC puede complicarse con Síndrome Urémico
Hemolítico (SUH o HUS) que puede llevar al paciente a la muerte por falla renal
aguda. E. coli O157:H7 es el serotipo
predominante en este grupo de EHEC.

Se sabe poco sobre las adhesinas, aunque se
ha reconocido un fenómeno de unión y borramiento similar a EPEC. Al igual que
EPEC se produce una fuerte adhesión y reorganización de actina. Una diferencia
seria que EHEC produce toxinas parecidas a la toxina Shiga, llamadas toxinas
similares a Shiga (SLTs). Es posible que la diarrea con sangre y HUS asociado a
EHEC sea debido a la producción de SLTs, aunque no seria raro la existencia de
otros factores intervinientes y aun no estudiados.
El gen que codifica para SLT se encuentra en
un fago temperado, lo que permitiría a otras cepas productoras de diarrea
adquirir SLT y dar una forma mucho más grave de enfermedad. Una característica importante
como factor de diseminación aunque no como factor de virulencia es la
posibilidad de la contaminación de carne durante la faena, que puede mezclarse
en la producción de hamburguesas, la mala cocción de estas en la preparación de
comidas rápidas, que ha llevado a la existencia de brotes en países desarrollados.
5. E.coli enteroinvasor (EIEC): Produce una enfermedad indistinguible de la disentería producida por Shigella.
Los pasos en la invasión y diseminación célula a célula parecen ser idénticos a
los de Shigella.
A diferencia de Shigella no produce toxina de Shiga. No se han descrito casos de
HUS en relación a estas cepas, probablemente en relación con la ausencia de
toxina Shiga. Al igual que Shigella muchos de los genes involucrados residen en un gran
plásmido de virulencia.

Las infecciones del tracto urinario
comienzan generalmente con la colonización de la uretra por cepas originarias
del colon previa colonización de la vagina. Una de las mayores defensas del
huésped es la acción lavadora de la orina. Las bacterias que no se pueden adherir
van a ser lavadas más rápidamente de la vejiga de lo que tardan en
multiplicarse.
Por otra parte las bacterias que adhieren
están más cerca de la mucosa y tienen mayores facilidades para provocar
respuesta inflamatoria. Numerosas adhesinas de E.coli uropatógeno
han sido estudiadas. Pili tipo 1 contribuyen a la colonización de la vagina y
parecen intervenir muy poco en el aparato urinario. La adhesina más importante,
sobre todo en cepas que causan infección renal es pili P.

Hay diversidad antigénica en estos pili pero
todos reconocen el mismo carbohidrato como receptor, globobiosa. Este azúcar se
encuentra unido a una ceramida anclada en la membrana de las células del huésped.
Estas cepas pueden poseer otras adhesinas que no son pili. Por ejemplo
adhesinas afimbriales (AFAI, AFAIII) o la adhesina Dr que reconocen las tres al
antígeno del grupo sanguíneo Dr como receptor. En general las cepas de E.coli uropatogénico
producen múltiples adhesinas por combinación de diferentes tipos de pili o
diferentes serotipos del mismo pili.
Esto podría permitir a las bacterias
adaptarse a diferentes superficies mucosas y ambientales, brindándole un mecanismo
de evasión de las defensas del hospedero. En cuanto a la respuesta
inflamatoria, hay evidencias de que LPS junto a pili P actúen sinérgicamente
provocando esta respuesta. Por otra parte algunas cepas uropatogénicas de E.coli producen
una exotoxina llamada hemolisina porque lisaba eritrocitos aunque luego se vio
que lisaba otras células.
Esta hemolisina (HlyA) pertenece a una gran familia de hemolisinas llamadas RTX. Todas ellas actúan creando poros
en las membranas celulares de los eucariotas. En el ratón las cepas que poseen
HlyA y pili P colonizan la vejiga, el riñón y matan dos tercios de los ratones testados,
por otra parte cepas isogénicas que producen solo pili P, colonizan pero no
causan daño renal ni muerte.
Las cepas que no poseen pili y no producen hemolisina
no colonizan. Al menos en el modelo animal la hemolisina media el daño renal. Los
genes que codifican para pili P están agrupados en el cromosoma. El conjunto
contiene genes para la subunidad mayor (pap
A), para las proteínas del tip (pap
E, F, G), para proteínas de procesamiento y ensamblado
(pap C, D, H, J, K) y proteínas reguladoras (pap B, I). Salvo el
gen I los demás forman un operón
transcripto desde un solo promotor.

Por otra parte los genes para hlyA también están agrupados y en
proximidad de los genes para pili. A las regiones que contienen los genes de
virulencia se las ha llamado Blocks de genes de virulencia.
MECANISMOS DE ACCIÓN DE E. COLI EN EL LUMEN INTESTINAL
Originalmente
descripta por Theodore Escherich en 1885 y llamada Bacterium coli
commune, hoy renombrada como Escherichia coli es uno de lo seres vivos
mas estudiados. Algunas cepas poseen diversos grados de patogenicidad, lo que
llevo a profundizar los mecanismos relacionados a ella. Otros patógenos se
adhieren a la célula huésped pegándose a proteínas preexistentes pero, en
Escherichia coli enteropatogénico se encontró un mecanismo diferente, ya que
manufactura e inyecta su propio receptor en la célula huésped para adherirse a
continuación. Las proteínas que intervienen se nombran con el prefijo Esp (de Enteropatgenic Escherichia coli Secreted Proteins).
La superficie de
las células epiteliales del intestino esta cubierta de microvellosidades,
extensiones de la célula que incrementan la superficie destinada a la absorción
de nutrientes. Escherichia coli (en color púrpura) se engancha a la
superficie de la célula epitelial del intestino (en marrón) por medio de los pili
(tetherlike pili). Los pili están constituidos por hebras de
largas proteínas filamentosas que pueden adherirse a las microvellosidades de
la superficie de las células intestinales.


Una vez en contacto con la bacteria desparecen las microvellosidades de una zona de la superficie celular, la bacteria entra en estrecho contacto con la superficie de la célula intestinal y comienza la siguiente fase del proceso de infección. La bacteria usa ahora un sistema especializado de inyección a fin de enviar algunas de sus propias proteínas al interior de la célula. Este sistema de inyección es fascinante y esta compuesto por numerosas proteínas. En este caso se esquematiza un sistema inyector Tipo III, el cual esta especializado para bombear cosas a otras células. La bacteria usa este sistema inyector como una jeringa e inyecta proteínas bacterianas en la célula, forzándola a cooperar con su propia infección.
Un tubo que hace las veces de aguja (en púrpura) denominado EspA, se proyecta desde la bacteria desde la bacteria a la superficie de la célula intestinal. Ahora dos proteínas (en verde) denominadas EspB y EspD viajan a través del tubo para formar una abertura en la membrana de la célula intestinal por medio de la cual proteínas bacterianas adicionales se mueven dentro de la célula configurando un poro. Cuando este proceso se completa la bacteria inyecta una proteína (en rojo) denominada Tir dentro de la célula.




Una vez dentro de
la célula la proteína Tir
se inserta en la membra celular y la "cabeza" de la misma se proyecta
mas allá de de superficie celular y se adhiere a a una proteína de la superficie bacteriana denominada intimina (las "ventosas"
azules). Ahora la bacteria esta adherida a la membrana de la célula intestinal,
y para la misma siguen los problemas. La proteína Tir resulta fosforilada por mecanismos
pertenecientes a la célula intestinal (esferas azules) y comienza el siguiente
paso: la formación del "pedestal".




La bacteria esta ahora adherida firmemente a la superficie celular por la interacción entre las proteínas Tir e intimina. Comienza ahora la formación del pedestal, un proceso notablemente activo. Otra proteína, perteneciente al citoesqueleto de la célula intestinal (en anaranjado) se adhiere a la porción de la proteína bacteriana Tir que se encuentra dentro de la célula.




Las imágenes corresponden a escenas
de la Animación Científica de B. Brett
Finlay, PhD realizada por Dennis Liu, Ph.D como Director y Eric Keller y
Satoshi Amagay, PhD como animadores.
Cuando numerosas
bacterias enteropatogénicas se han adherido comienzan los síntomas de la
infección (diarrea). Se cree que algunas de las proteínas inyectadas por Escherichia
coli, como la EspB ,
interfieren los mecanismos de señales de la célula huésped lo cual lleva,
eventualmente, al desarrollo de lesiones ultraestructurales y a la diarrea
secretoria observada durante la infección.
ESCHERICHIA COLI EN EL MARCO DE LA SEGURIDAD ALIMENTARIA
Su
comportamiento y su difusión, al ser un microorganismo intestinal, se asocian a
Salmonella, por lo que las medidas preventivas básicas a tomar son
similares para ambos casos. Esto es, debe extremarse la higiene personal, sobre
todo en el caso de ser portadores del patógeno, y evitar el consumo de
alimentos crudos o poco o deficientemente cocinados.
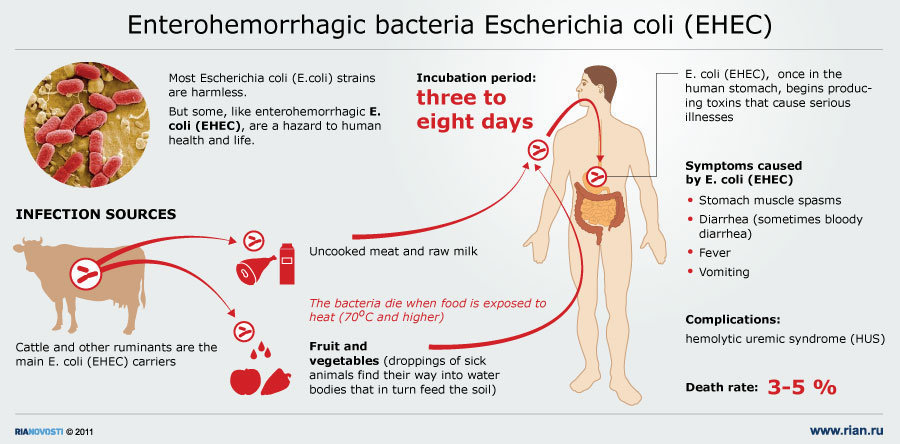
Su detección, por otra parte, es relativamente simple por los sistemas de control rutinario de cualquier laboratorio. Aunque la normativa actualmente vigente no lo exija, desde distintos sectores se ha venido insistiendo en la necesidad de aplicar estas rutinas con el fin de limitar un riesgo considerado evitable. El control se plantea de forma especial para con los alimentos crudos como para la carne y sus derivados, el pescado y sus derivados, y los vegetales.
La simple presencia de este microorganismo, o un recuento superior a 100 ufc/g o ml indicará una contaminación fecal con el consiguiente riesgo de que existan cepas patógenas. Aunque en general las enteritis cursan de forma característica (colitis hemorrágica afebril), la causada por la bacteria verotoxigénica da lugar a manifestaciones variables que van de formas muy leves a formas graves con sangre (colitis hemorrágica). Se ha podido constatar que la fiebre es relativamente frecuente en los casos de enteritis causada por la variante O157:H7, así como la complicación con el Síndrome Hemolítico Urémico. Los mecanismos por los cuales se producen no se conocen con precisión.

La infección por E. coli verotoxigénica parece ser de distribución universal, aunque irregular, pero su prevalencia solamente se conoce con cierto detalle en los Estados Unidos, Canadá, Argentina y Europa Occidental, ya que en el resto de países no ha sido estudiada sistemáticamente. Diversos autores han estudiado en España la frecuencia de E. coli O157:H7 como causante de diarrea y se ha podido demostrar que ésta es muy baja, probablemente entre el 0,1 y 1% de las diarreas estudiadas. Normalmente se detectan casos esporádicos aunque no son raros los brotes epidémicos y en general, no obstante, su número y extensión son limitados.

La enfermedad se transmite por vía feco – oral y el vehículo más frecuente de infección humana es la carne de bovino, fundamentalmente las hamburguesas poco cocinadas. También se ha documen-tado la infección vehiculada por otros alimentos como carne de pavo, salames, leche, yogur, mayo- nesa, ensaladas, vegetales crudos y agua. Los brotes epidémicos son frecuentes en diversos países como Estados Unidos, Reino Unido, Australia, Argentina y Japón, entre otros. La transmisión de persona a persona también ha sido demostrada y la dosis infectante mínima se estima alrededor de las 100 bacterias. Las distintas formas de Escherichia coli suelen ser resistentes a las temperaturas extremas y a los ácidos débiles.

Los bóvidos parecen constituir el principal reservorio de E. coli O157:H7, encontrado con diferen-tes prevalencias que oscilan, en animales sanos, entre el 7% y el 30% de los casos estudiados. Pa-rece que estas cepas no son patogénicas para los animales, aunque algunos investigadores las en-cuentran con más frecuencia en aquellos que tienen diarrea. La prevalencia de otros serotipos de E. coli verotoxigénicos en los animales se desconoce, aunque hay informes de su aislamiento en bó-vidos, óvidos, cabras, perros y gatos. Desde 1986, diversos grupos han efectuado estudios prospectivos en nuestro país, que muestran una incidencia muy baja de E. coli verotoxigénica inferior al 0,3% de los pacientes estudiados.

Con respecto a los
vegetales que se cultivan abonándolos con estiércol animal, hasta hoy se
consideraba que, después de la recolección, había que proceder al lavado de las
verduras frescas, normalmente con la incorporación al agua de desinfectantes,
como el hipoclorito de sodio (lejía) o la mezcla de peróxido de hidrógeno y
ácido peracético. Sin embargo, las autoridades sanitarias de EEUU han
comprobado que los lavados prescritos se han mostrado totalmente ineficaces contra
este microorganismo.




BUENAS TARDES. POR FAVOR SI ME PUEDE AYUDAR CON MAYOR INFORMACION SON TEMAS DE MUCHA RELEVANCIA. AL CORREO ( freire.mariela@gmail.com)
ResponderBorrarGRACIAS POR SU COMENTARIO MARIELA. LE ESCRIBÍ PARA SABER QUE NECESITA ESPECÍFICAMENTE. SALUDOS CORDIALES.
ResponderBorrar